近两年来,多项研究发现染色质中的新生RNA上含有m6A修饰,说明这些m6A修饰伴随着转录产生。真核生物的转录包括转录起始、暂停与释放、延伸和终止等多个步骤。早期的转录暂停以及暂停后命运选择(继续延伸还是降解)是高等生物基因转录的限速步骤,而m6A修饰在此过程中的研究还很少。
mRNA m6A修饰的功能近十年来得到了广泛的关注。mRNA上的m6A修饰主要由METTL3/METTL14/WTAP复合物催化,参与了mRNA的剪接、转运、降解、翻译等过程。但这些研究主要集中在m6A对于转录后mRNA的命运调控,关于新生RNA上的m6A功能的研究直到近两年才逐渐展开。
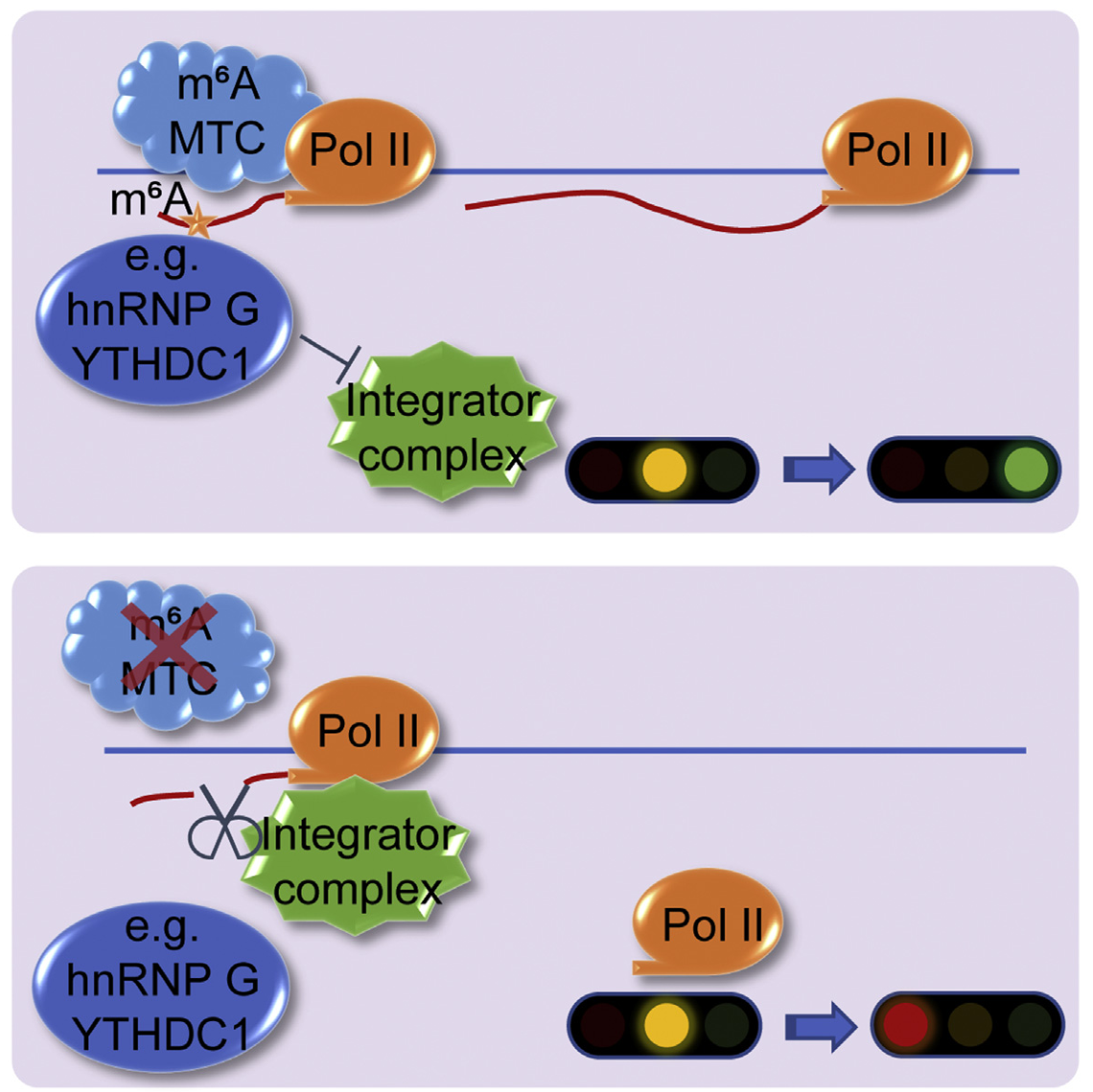
2022年2月25日,复旦大学生物医学研究院研究员沈宏杰、蓝斐联合牛津大学Ludwig肿瘤研究所教授Yang Shi和哈佛医学院教授Karen Adelman合作研究成果,以“Dynamic Control of Chromatin-associated m6A Methylation Regulates Nascent RNA Synthesis”为题,在线发表于Molecular Cell杂志。该研究发现伴随转录产生的新生RNA m6A修饰可促进转录,该功能通过抑制Integrator复合物招募而实现,揭示了m6A修饰对于转录调控的重要作用。

研究者发现在乳腺癌MCF-7细胞、人胚胎肾HEK293T细胞以及小鼠神经干细胞(Neural Stem Cell)中,METTL3、METTL14和WTAP主要结合在基因启动子和增强子上的转录起始位点附近。研究者发现METTL3可以催化染色质结合的新生RNA上的m6A修饰,不同于成熟mRNA的m6A修饰主要位于3'UTR,新生RNA的5'UTR 也含有m6A修饰,这些m6A修饰可以被hnRNP G和YTHDC1识别,从而阻止Integrator复合物剪切新生RNA,促进了新生RNA转录。
值得一提的是沈宏杰课题组在2021年的一篇研究还发现小鼠胚胎干细胞中,METTL3主要结合在异染色质区域,通过组蛋白H3K9me3甲基化,抑制内源性逆转录病毒IAPEz元件的转录(Nature,2021)。而随着小鼠胚胎干细胞分化到拟胚体(Embryoid Body,EB)时期,METTL3在染色质上的分布则迁移到了启动子区域,提示METTL3在退出自我更新状态后更多的参与活化区域转录起始调控,(Nature,2021,复旦网站链接:https://news.fudan.edu.cn/2021/0129/c4a107857/page.htm)。
原文链接:https://doi.org/10.1016/j.molcel.2022.02.006




